{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
TRPC对被动致敏的人气道平滑肌细胞的增殖及下游基因表达的作用
引用本文
闫文丽, 张晓宇, 唐学义. TRPC对被动致敏的人气道平滑肌细胞的增殖及下游基因表达的作用. 循证医学, 2021,21(5): 283-288.
YAN Wen-li, ZHANG Xiao-yu, TANG Xue-yi. Effects of TRPC on the Proliferation and Expression of Downstream Genes in Passively Sensitized Human Airway Smooth Muscle Cells. Journal of Evidence-Based Medicine,2021,21(5): 283-288.
DOI:10.12019/j.issn.1671-5144.2021.05.007
Permissions
YAN Wen-li, ZHANG Xiao-yu, TANG Xue-yi. Effects of TRPC on the Proliferation and Expression of Downstream Genes in Passively Sensitized Human Airway Smooth Muscle Cells. Journal of Evidence-Based Medicine,2021,21(5): 283-288.
DOI:10.12019/j.issn.1671-5144.2021.05.007
Copyright©2021, 《循证医学》编辑部
《循证医学》杂志 版权所有
TRPC对被动致敏的人气道平滑肌细胞的增殖及下游基因表达的作用
闫文丽(1975-),女,河南安阳人,副主任医师, 医学学士,主要研究方向为呼吸道感染性疾病。
摘要
目的 探究经典瞬时感受器电位通道(transient receptor potential channels,TRPC)对被动致敏的人气道平滑肌细胞(human airway smooth muscle cells,HASMC)的增殖及其表达和作用。方法 本实验设置分组情况如下:实验组TRPC6阻断剂SKF96365处理的HASMC细胞;TRPC6 siRNA转染的HASMC细胞;对照组TRPC6野生型的HASMC细胞。对不同处理的细胞利用实时荧光定量聚合酶链反应、western blot、细胞计数以及氚胸腺嘧啶核苷掺入试验检测其增殖及其表达和作用。结果 利用western blot检测TRPC6蛋白发现TRPC6阻断剂SKF96365和TRPC6 siRNA转染均能够显著减少HASMC中TRPC6的表达( P<0.05)。氚胸腺嘧啶核苷(tritium thymine nucleoside,3H-TdR)掺入实验对TRPC6对HASMC细胞增殖的检测发现10 μmol/L的SKF96365处理72 h或TRPC6 siRNA转染细胞48h后,3H-TdR标记的细胞数量显著减少( P<0.05)。细胞计数实验发现,TRPC6阻断剂SKF96365使TRPC6基因表达减少后,细胞数目随着SKF96365浓度的升高而减少( P<0.01),在单一浓度10 μmol/L的作用下,细胞数目随着SKF96365处理时间的延长增长速度缓慢( P<0.01),同时通过TRPC6 siRNA转染使TRPC6敲除后细胞增殖明显减少。TRPC6阻断剂SKF96365和TRPC6 siRNA转染抑制了TRPC6的表达后,均能够显著降低HASMC中PI3K、AKT、p-AKT、mTOR的表达( P<0.05)。结论 TRPC6通过PI3K/Akt/mTOR通路对被动致敏的HASMC的增殖及其表达具有显著的抑制作用,因此本研究为哮喘病的治疗提供新的参考依据。
关键词:
哮喘; TRPC6; HASMC; 细胞增殖
中图分类号:R56;R329
文献标识码:A
收稿日期: 2020-11-13
Effects of TRPC on the Proliferation and Expression of Downstream Genes in Passively Sensitized Human Airway Smooth Muscle Cells
Abstract
Objective To explore the proliferation, expression and effect of TRPC on passively sensitized human airway smooth muscle cells (HASMC).Methods HASMC cells treated with TRPC6 blocker SKF96365 in the experimental group were grouped as follows: TRPC6 siRNA transfected HASMC cells; Control group TRPC6 wild type HASMC cells. The proliferation, expression and function of tritium thymine nucleoside were detected by real-time fluorescence quantitative polymerase chain reaction, western blot, cell count and tritium thymine nucleoside (3H-TdR) addition test.Results Western blot analysis of TRPC6 protein showed that TRPC6 blocker SKF96365 and TRPC6 siRNA transfection could significantly reduced the expression of TRPC6 in HASMC ( P<0.05). When 3H-TdR was added to HASMC, it was found that the number of 3H-TdR labeled cells decreased significantly after 10 mol/L SKF96365 treatment for 72 h or TRPC6 siRNA transfection for 48 h ( P<0.05). Cell count experiment found that TRPC6 blocker SKF96365 reduced TRPC6 gene expression, the number of cells with the rising of SKF96365 concentration reduced ( P<0.01), in a single concentrations under the action of 10 μmol/L, cell number as SKF96365 processing time slowed growth rate ( P<0.01), and at the same time through TRPC6 siRNA transfection cell proliferation significantly reduced after the TRPC6 knocked out. After TRPC6 blocking agents SKF96365 and TRPC6 siRNA were transfected, the expression of PI3K, Akt, p-Akt and mTOR in HASMC were significantly reduced.Conclusions TRPC6 had a significant inhibitory effect on the proliferation and expression of downstream genes in passively sensitized HASMC, through PI3K/Akt/mTOR pathway. Therefore, this study provided a new reference for the treatment of asthma.
Key words:
asthma; TRPC6; HASMC; cell proliferation
哮喘是人类较为常见的多基因遗传病, 该疾病的发生受到多种细胞和组织的调控, 遗传因素和环境因素均会对哮喘造成部分影响[1, 2]。支气管哮喘属于一种慢性炎症性疾病, 基本特征为气道高反应性和可逆性气流阻塞, 其中气道重塑是哮喘的主要特征之一。哮喘气道重塑时会发生多种现象, 其中最典型的为气道结构的改变, 主要病理性改变为气道平滑肌细胞(airway smooth muscle cells, ASMCs)的肥大和增生。目前, 对于难治性哮喘病的治疗, 抑制ASMCs的过度增殖为最佳途径[3, 4, 5]。对相关文献查找发现, 针对气道平滑肌细胞增殖调控机制尚不明确。目前, 已知的数据表明, 钙离子在细胞增殖调控中发挥重要作用, 而经典瞬时感受器电位通道(transient receptor potential channels, TRPC)是控制细胞内钙离子水平的重要通道。TRPC家族成员众多, 包括TRPC1、TRPC2、TRPC3、TRPC4、TRPC5、TRPC6、TRPC7七种, 可严格调节钙离子的内流和外流。已有文献报道, TRPC6对大鼠气道平滑肌细胞的增殖具有重要调节作用[6, 7, 8]。因此, 本文主要针对TRPC家族成员TRPC6对被动致敏的人气道平滑肌细胞(human airway smooth muscle cells, HASMC)的增殖及其表达和作用进行探究, 为哮喘病的治疗提供新的参考依据。
1 材料与方法
1.1 主要材料及分组
HASMC购买于美国典型培养物保藏中心(American Type Culture Collection, ATCC), 将HASMC培养于含10%哮喘患者血清(IgE总水平> 200 U/mL)的DMEM培养液中被动致敏24 h。本实验设置分组情况如下:实验组TRPC6阻断剂SKF96365处理的HASMC细胞; TRPC6 siRNA转染的HASMC细胞; TRPC6野生型的HASMC细胞。本研究主要实验材料如下:SYBR Green荧光染料, 兔多克隆TRPC6抗体, 山羊抗兔多克隆抗体, TRPC6上、下游引物, GAPDH上、下游引物, RIPA细胞组织裂解液, TRPC6阻断剂SKF96365, TRPC6 siRNA, 氚胸腺嘧啶核苷。
1.2 方法
1.2.1 实时荧光定量聚合酶链反应
利用实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction, RT-PCR)检测HASMC中TRPC6 mRNA的含量:(1)细胞总RNA的提取:总RNA提取过程需严格根据Trizol组织或细胞总RNA提取方法完成实验操作。(2)逆转录合成cDNA:按照逆转录试剂盒说明书各步骤严格操作, 进行cDNA的合成。荧光染料选择SYBR Green。根据引物设计原则设计的TRPC6引物序列如下:上游引物5′ -GCCAATGAGCATCTGGAAAT-3′ , 下游引物5′ -TGGAGTCACATCATGGGAGA-3′ 。内参基因磷酸甘油醛脱氢酶GAPDH引物序列如下:上游引物5′ -ATGGGGAAGGTGAAGGTCG-3′ , 下游引物5′ -GGGGTCATTGATGGCAACAATA-3′ 。(3)RT-PCR步骤:本实验选择的反应体系为40 μ L, 上游引物0.5 μ L, 下游引物0.5 μ L, DNA模板溶液1 μ L, 10× PCR缓冲液5 μ L, 浓度为2.5 mmol/L的dNTP 4 μ L, 浓度为25 mmol/L的MgCl2 4 μ L, 浓度为5× 106U/L的Taq DNA 聚合酶0.5 μ L, SYBR Green荧光染料1 μ L, 最后用去除RNase的无菌水将体系补齐。上机检测时需将样品与阳性对照和空白对照同时检测, RT-PCR反应条件设置为95℃ 3 min, 92℃ 1 min, 60℃ 45 s, 72℃ 45 s, 一共循环35次。
1.2.2 Western blot 检测TRPC6蛋白的表达
利用western blot检测TRPC6蛋白在HASMC中表达:(1)总蛋白的提取:本研究通过RIPA细胞组织裂解液对细胞的总蛋白进行提取。蛋白裂解后, 加入适量的SDS loading buffer于100℃金属浴中煮样10 min, 置于冰上冷却后可进行SDS-PAGE电泳分离。(2)SDS-PAGE电泳:利用SDS-PAGE电泳将总蛋白进行分离, 浓缩胶电泳时电压设置为70~80 V, 待样品电泳至分离胶时, 可将电压调至120~130 V, 直至将蛋白样品彻底分离。(3)转膜:将SDS-PAGE凝胶上的蛋白样品转移至硝酸纤维素膜上, 该过程采用湿转方式, 转膜参数设置为恒定电流300 mA, 时间为80 min。(4)封闭:将硝酸纤维素膜用5%的脱脂牛奶封闭, 目的是封闭非特异性抗原, 避免过多的非特异性显色。(5)一抗:根据marker剪下目标蛋白分子量大小的条带, 利用兔多克隆TRPC6抗体进行一抗的孵育, 条件为4℃过夜。(6)二抗:一抗孵育完毕后将条带用PBST缓冲液清洗3~5次, 然后采用含有ECL发光基团的山羊抗兔多克隆抗体即二抗进行孵育, 条件为室温1~2 h。(7)显色:将条带滴加适量ECL显影液后置于显影仪中进行条带显色。用Image J分析软件对目标条带进行分析, TRPC6的相对表达水平以TRPC6与内参蛋白GAPDH的比值计量。
1.2.3 细胞计数检测HASMC的增殖情况
将被动致敏的HASMC进行实验分组, 分为对照组和研究组, 每组含有相同的细胞个数。其中研究组使用不同浓度的TRPC6阻断剂SKF96365处理72 h, 浓度梯度选择5、10、15、20 μ mol/L, 该药物配制时的溶剂为二甲基亚砜(dimethyl sulfoxide, DMSO), 因此对照组使用相同体积的DMSO处理72 h。在药物处理的不同时间点(0 h、24 h、48 h、72 h、96 h)对每种条件下的细胞进行计数, 全部完成后制作细胞生长曲线。
1.2.4 TRPC6 siRNA的转染和氚胸腺嘧啶核苷掺入试验
除了利用TRPC6抑制剂来减少TRPC6的表达进而检测其对细胞增殖的影响外, 本研究还通过转染TRPC6 siRNA来沉默TRPC6。根据相关引物设计原则得出TRPC6 siRNA的正义链序列为5′ -TTCCCGCCATGAGCCAGA-3′ , 反义链序列为5′ -TCTGGCTCATGGCGGGAA-3′ 。根据碧云天的脂质体lipofectamine 3000转染试剂盒将TRPC6 siRNA严格根据操作流程转染HASMC, 转染时间为48 h, 然后通过western blot检测TRPC6蛋白的表达并利用氚胸腺嘧啶核苷掺入实验来检测其对细胞增殖的影响。
1.3 统计学方法
数据资料均通过SPSS 22.0软件完成数据统计学分析, 以P< 0.05表示对比差异存在统计学意义。计量数据通过“ x ± s” 形式表示, 比较行t检验; 计数数据比较行χ 2检验。
2 结果
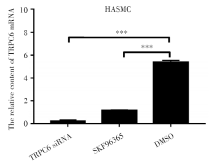
2.1 RT-PCR检测实验组和对照组中TRPC6 mRNA表达情况
RT-PCR检测实验组和对照组中TRPC6 mRNA表达情况如图1所示, TRPC6 mRNA在HASMC中的表达量与SKF96365处理和TRPC6 siRNA转染相比显著升高(P< 0.001), 差异具有统计学意义。
 | 图1 TRPC6 mRNA在HASMC中的相对表达情况 注:RT-PCR检测HASMC的TRPC6 mRNA的相对表达。* * * P< 0.001。每个条形图代表三个独立实验的平均值± SDFig.1 The expression of TRPC6 mRNA in HASMC Note:Relative expression of TRPC6 mRNA in HASMC as determined by RT-PCR. * * * P< 0.001. Each bar represents the mean ± SD of three independent experiments |
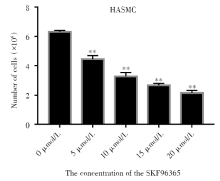
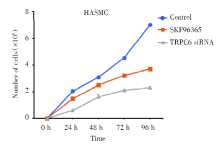
2.2 细胞计数检测TRPC6对HASMC细胞增殖的影响
利用细胞计数的方式对不同处理条件下的HASMC的增殖情况进行测定, 结果如图2、图3所示, TRPC6阻断剂SKF96365使TRPC6基因表达减少后, 细胞数目随着SKF96365浓度的升高而减少(P< 0.01), 在单一浓度10 μ mol/L的作用下, 细胞数目随着SKF96365处理时间的延长增长速度缓慢(P< 0.01), 同时通过TRPC6 siRNA转染使TRPC6敲除后细胞增殖明显减少, 表明TRPC6能够抑制被动致敏的HASMC的增殖和表达。
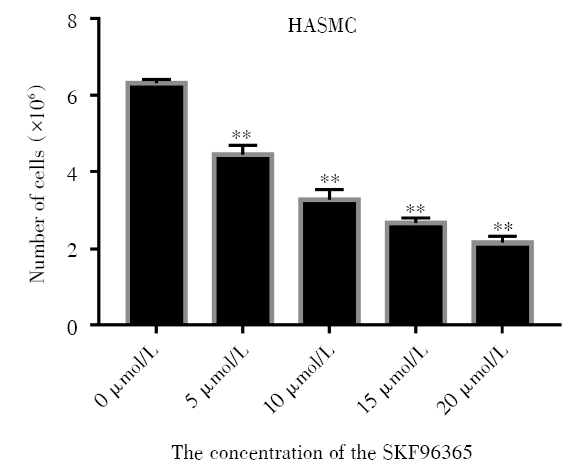 | 图2 SKF96365不同浓度对细胞增殖的影响 注:将HASMC培养于含不同浓度(0、5、10、15、20 μ mol/L) SKF96365的培养基中, 72 h后计数细胞数量。* * P< 0.01。每个条形图代表三个独立实验的平均值 ± SDFig.2 The effect of SKF96365 at different concentrations on HASMC proliferation Note: HASMC were cultured in medium containing different concentrations(0, 5, 10, 15, 20 μ mol/L) of SKF96365, then cells were counted after 72 h. * * P< 0.01. Each bar represents the mean ± SD of three independent experiments |
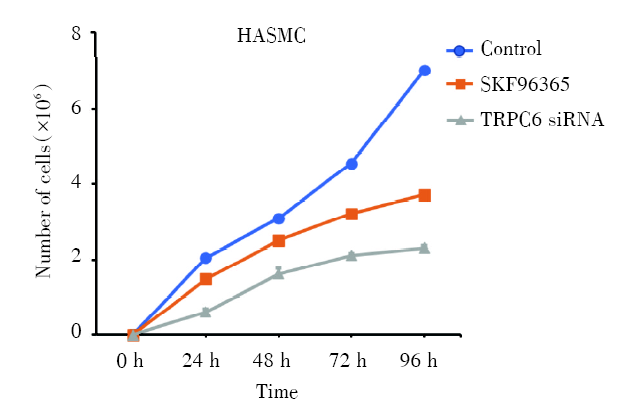 | 图3 SKF96365、TRPC6 siRNA对细胞增殖的影响 注:分别在不同时间点(0 h、24 h、48 h、72 h、96 h)对正常HASMC、10 μ mol/LSKF96365处理HASMC和TRPC6 siRNA转染HASMC进行计数。每个条形图代表三个独立实验的平均值 ± SDFig.3 The effect of SKF96365 and TRPC6 siRNA on HASMC proliferation Note: Normal HASMC, HASMC treated with 10 μ mol/LSKF96365 and HASMC transfected with TRPC6 siRNA were counted at different time points (0 h, 24 h, 48 h, 72 h, 96 h). Each bar represents the mean± SD of three independent experiments |
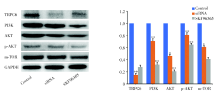
2.3 TRPC6通过PI3K/Akt/mTOR通路影响HASMC增殖
Western blot检测TRPC6蛋白的表达结果如图4所示, TRPC6阻断剂SKF96365和TRPC6 siRNA转染均能够显著减少HASMC中TRPC6的表达。为了明确TRPC6影响HASMC增殖所涉及的相关分子机制, 应用western blot检测了对照组、siRNA敲减TRPC6组以及TRPC6阻断剂SKF96365组中磷脂酰肌醇3-激酶/蛋白激酶B/雷帕霉素靶蛋白(phosphatidylin-ositol-3-kinase/protein kinase B/mammalian target of rapamycin, PI3K/Akt/mTOR)信号通路的表达情况, PI3K/Akt/mTOR信号通路能够促进包括人平滑肌在内的多种类型细胞增殖, 结果如图4所示, TRPC6阻断剂SKF96365和TRPC6 siRNA转染抑制了TRPC6的表达后, 均能够显著降低HASMC中PI3K、AKT、p-AKT、mTOR的表达。
 | 图4 SKF96365和TRPC6 siRNA处理后TRPC6蛋白及PI3K/Akt/mTOR通路分子的表达情况 注:TRPC6蛋白和PI3K/Akt/mTOR在正常HASMC、SKF96365处理HASMC和转染TRPC6 siRNA的HASMC中的相对表达及western blot定量分析。* P< 0.05; * * P< 0.01。每个条形图代表三个独立实验的平均值 ± SDFig.4 The expression of TRPC6 protein and PI3K/Akt/mTOR in normal HASMC, HASMC treated with SKF96365 and HASMC transfected with TRPC6 siRNA Note:Relative expression and quantitative analysis of western blot of TRPC6 protein and PI3K/Akt/mTOR in normal HASMC, HASMC treated with SKF96365 and HASMC transfected with TRPC6 siRNA. * P< 0.05; * * P< 0.01. Each bar represents the mean ± SD of three independent experiments |
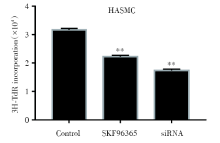
2.4 氚胸腺嘧啶核苷掺入实验检测TRPC6对HASMC增殖的影响
氚胸腺嘧啶核苷(tritium thymine nucleoside, 3H-TdR)掺入实验对TRPC6对HASMC细胞增殖的检测结果如图5所示, 10 μ mol/L的SKF96365处理72 h或TRPC6 siRNA转染细胞48 h后, 3H-TdR标记的细胞数量显著减少(P< 0.01), 差异具有统计学意义。
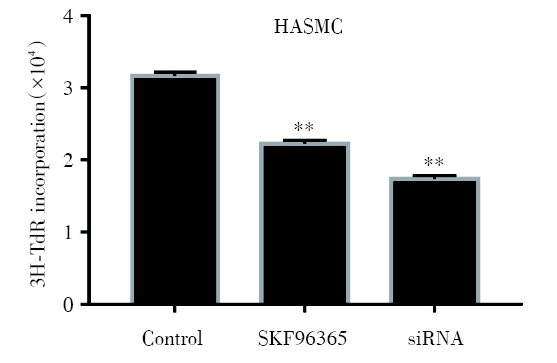 | 图5 SKF96365和TRPC6 siRNA处理后3H-TdR掺入情况 555注:将3H-TdR加入正常HASMC、SKF96365处理HASMC和TRPC6 siRNA转染HASMC中, 10 mol/L SKF96365处理72 h和TRPC6 siRNA转染48 h后检测3H-TdR标记细胞数。* * P< 0.01。每个条形图代表三个独立实验的平均值 ± SDFig.5 The content of 3H-TdR in normal HASMC, HASMC treated with SKF96365 and HASMC transfected with TRPC6 siRNA Note:Tritium thymine nucleoside was added to normal HASMC, HASMC treated with SKF96365 and HASMC transfected with TRPC6 siRNA, then the number of 3H-TdR labeled cells was detected after 10 mol/L SKF96365 treatment for 72 h and TRPC6 siRNA transfection for 48 h. * * P< 0.01. Each bar represents the mean ± SD of three independent experiments |
3 结论
TRPC是TRP离子通道蛋白超家族成员之一, 目前已公认的TRPC通道存在7种, 各成员之间相互协调, 共同控制着人体的相关功能[9, 10, 11]。TRPC家族成员可分为TRPC1、TRPC2、TRPC3、TRPC4、TRPC5、TRPC6、TRPC7, 虽然以上离子通道同属于一个家族, 但其通道激活途径、一般属性以及生理功能并不一致。有文献报道, TRPC通道调节神经元发育, 与神经元细胞的增殖分化密切相关, 同时其对神经分泌和长期电位具有显著调节作用[12, 13, 14]。根据TRPC家族成员间的结构、功能以及激活方式将其进行分类, 可分为钙离子耗竭激活的由TRPC1、TRPC4、TRPC5组成的第一组, 受体刺激激活的由TRPC3、TRPC6、TRPC7组成的第二组。TRPC通道蛋白的主要作用包括在细胞中利用异构分子之间相互作用进而形成同源四聚体或异源四聚体对钙离子浓度进行调节, 从而保持相对稳定状态; 在生理状态下, TRPC通道的激活或抑制也调节着磷脂酶C介导的信号转导通路; 与神经元的基本生理作用密切相关; 另外, 该通道功能的异常与老年痴呆症、小脑性共济失调以及帕金森症等疾病的发生具有部分相关性[15, 16, 17]。
本研究主要探讨的TRPC6, 主要存在部位为嗅觉上皮细胞、视网膜神经节细胞以及脑的部分区域, 例如海马区、小脑区、皮质区等。TRPC6与多种细胞类型细胞增殖相关, 能够促进肾小管上皮细胞、肺癌细胞、肺动脉平滑肌细胞等的增殖[18, 19, 20]。已有文献报道TRPC6对大鼠气道平滑肌细胞的增殖具有重要调节作用[6, 7, 8], 而TRPC6对被动致敏的HASMC的增殖调节作用和相关的机制尚未被研究。
为了验证抑制TRPC6的表达, 是否会抑制HASMC的增殖, 本研究首先利用TRPC6阻断剂SKF96365和TRPC6 siRNA转染两种方式干扰TRPC6的表达, 均能够显著减少HASMC中TRPC6的表达。利用细胞计数实验将对照组、TRPC6阻断剂SKF96365组处理后的HASMC细胞数目进行检测发现, TRPC6阻断剂SKF96365使TRPC6基因表达减少后, 细胞数目有所减少, 且细胞数减少的程度与SKF96365浓度的升高呈正相关(P< 0.01), 具有浓度依赖。选择单一浓度10 μ mol/L作用HASMC, 细胞数目则随着SKF96365处理时间的延长增长速度缓慢(P< 0.01)。同时, 通过TRPC6 siRNA转染, 同样使TRPC6敲除后HASMC细胞增殖明显受到抑制, 且对于细胞增殖的抑制效果要强于TRPC6阻断剂处理, 推测原因可能是小干扰RNA的敲除作用对于TRPC6的抑制程度更强, TRPC6阻断剂加入细胞后, 并不能保证阻断剂能够持续维持有效的作用浓度, 因而对TRPC6的抑制作用不及siRNA, 从而抑制细胞增殖能力不及siRNA转染。
本研究也利用氚胸腺嘧啶核苷酸掺入实验, 将对照组、TRPC6阻断剂SKF96365组和TRPC6 siRNA转染组处理后的HASMC细胞增殖情况进行检测, 发现10 μ mol/L的SKF96365处理72 h或TRPC6 siRNA转染细胞48 h后, 3H-TdR标记的细胞数量显著减少(P< 0.05)。以上实验均表明, 抑制TRPC6能够显著抑制HASMC增殖。
明确了TRPC6正向调节HASMC增殖过程, 本研究继续对该过程所涉及的相关通路及分子进行验证。通过文献检索得知, TRPC6调节细胞增殖过程与P13K/Akt/mTOR信号通路相关, TRPC6通道介导的PC12细胞和海马神经元突起生长涉及PI3K信号通路的激活[21]; 在基因敲除TRPC6-/-小鼠中Akt/mTOR通路的激活也下调[22]。P13K/Akt/mTOR信号通路的功能障碍与许多人类疾病有关, 而各种钙依赖性信号级联通过激酶和磷酸酶直接或间接地受到P13K/Akt信号通路的影响。本研究利用western blot检测了上述三种细胞中PI3K/Akt/mTOR信号通路的表达情况, 结果显示TRPC6阻断剂SKF96365和TRPC6 siRNA转染抑制了TRPC6的表达后, 均能够显著降低HASMC中PI3K、Akt、p-Akt、mTOR的表达。由此说明, TRPC6对于HASMC细胞增殖的抑制作用, 与其抑制P13K/Akt/mTOR信号通路相关。
综上, TRPC6能够通过抑制P13K/Akt/mTOR信号通路从而抑制被动致敏的HASMC的增殖和表达, 为哮喘病的治疗和相关研究提供新的参考依据。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|